![[Logo] Instituto Superior
Técnico](img/wwwist.gif)
Aviso:Se está a ler esta mensagem, provavelmente, o browser que utiliza não é compatívelcom os "standards" recomendados pela W3C.Sugerimos vivamente que actualize o seu browser para ter uma melhor experiênciade utilização deste "website".Mais informações em webstandards.org.
Warning: If you are reading this message, probably, your browser is not compliant with the standards recommended by the W3C. We suggest that you upgrade your browser to enjoy a better user experience of this website.More information on webstandards.org.

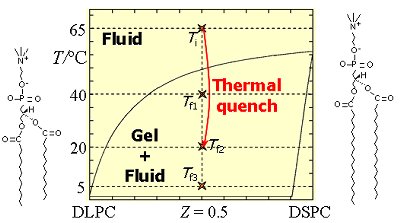
Membrane rafts and lipid domainsIt has been long realized that model systems of membranes prepared with phospholipid mixtures with different main transition temperatures (Tm) can exhibit lateral phase coexistence of gel/fluid phases. Depending on the ideality of the mixture, gel-fluid and even gel-gel and fluid-fluid phase separation can occur. Another group of lipid mixtures that have been intensely used are phospholipid (mainly PC)/cholesterol (chol) binary systems, for which liquid ordered (ld)/liquid disordered (lo) phase separation may occur. These mixtures' properties and interaction with peptides and proteins is of importance due to the high abundance of chol in mammalian plasma membranes. The fact that phase separation occurs for mixtures of lipids coexisting in cell membranes under conditions close to physiological made the detection and characterization of this kind of lateral heterogeneity of considerable interest in the biophysical and biochemical communities. The composition-temperature phase diagrams, at constant pressure (and ionic strength, etc.), are a convenient way to represent this type of behavior for each pair of phospholipids. However, they do not present a complete description of the phase separation phenomenon, because, at variance with common 3D phase separation in binary mixtures, this process may not go to completion, but lead to the formation of smaller portions (domains) of the least abundant phase, dispersed in the other phase. Lipid rafts are a special type of membrane domains. They were postulated in 1997, and are thought to consist of sphingolipid/chol- enriched and (unsaturated) phosphatidylcholine (PC)-depleted domains. They are involved in important cellular processes such as sorting and trafficking of lipids and proteins, and signal transduction. The simplest model for rafts is a mixture of a high Tm lipid with a low Tm lipid and chol. The rafts would correspond to the lo domains enriched in high Tm lipid and chol, dispersed in a ld matrix enriched in low Tm lipid. Apparently, rafts in a rest state in cell membranes are in general <100 nm. In our group, the properties of several binary and ternary lipid mixtures (vesicles) have been studied by fluorescence spectroscopy. Concerning the binary mixtures (for which the phase diagram was previously known) the size of domains was estimated for mixtures of two phospholipids with gel/fluid phase separation (stemming from a large difference of acyl chain length in the two constituents). The slow kinetics of phase separation after sudden cooling from a temperature above the Tm of both lipids was also studied (Figures 1 and 2).
PC/chol binary mixtures with ld/lo phase separation are usually more difficult to study, because the two coexisting phases are more similar than for gel/fluid separation. However, the use of fluorescence techniques (namely FRET) made it possible to study the domains for different experimental conditions (composition, temperature).
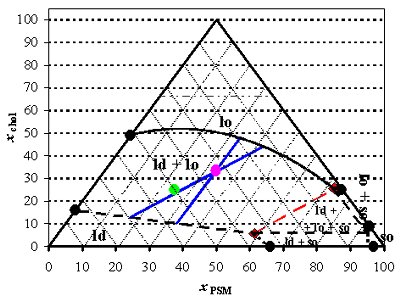
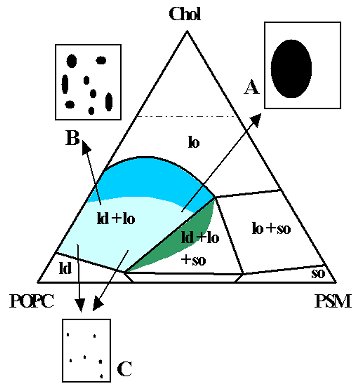
Recently, we published the phase diagram for the raft model ternary system sphingomyelin (SM)/PC/chol, obtained by fluorescence methods (Figure 3). The diagram gives the composition and boundaries of the rafts, explains apparently conflicting literature results, and, because the direction of some tie-lines was also determined, allows carrying out for the first time thermodynamically consistent studies on that ternary system, namely the interaction of other molecules with raft model membranes. The knowledge of the tie-line also allowed carrying out a FRET study similar to those carried out in the binary systems (Figure 4). Other ternary systems of considerable biological relevance are currently under study.
Ceramide (Cer), a biosynthetic precursor of sphingomyelin that can be also generated by the action of sphingomyelinase in cell membranes, has recently emerged as a key molecule in the modulation of several cellular processes such as apoptosis and stress-signalling cascades, probably by inducing the formation of large raft platforms. Cer is considered to act as a second messenger in various signalling pathways and its action is commonly ascribed to changes in the membrane physical properties after Cer generation. Increase in membrane order, microdomain formation (gel-fluid phase separation) and formation of non-lamellar lipid phases are frequently reported for this lipid. Both binary (Cer/PC) and ternary (Cer/PC/chol) mixtures are currently being studied. |
||||||||||||